Related Documentation:
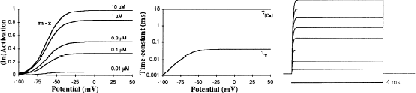
In their single-channel study of K+ channels in Purkinje cells, [2] describe 2 Ca2+-activated K+ channels. One is a 134 pS (symmetrical K+ conditions) channel, which they identify as the K2 channel. These authors report that this channel composes ~ 13 % of the total Ca2+-activated K+ conductance, activates at lower potentials than the BK channel, and is blocked by TEA. However, [2] does not contain enough data to model this channel. The K2 channel is not a small K channel (SK) channel, because SK channels have a lower slope conductance (10–20 pS), are not voltage sensitive [3, 4] and are blocked by apamin [5]. In rat brain synaptosomal membranes several types of Ca2+-activated K+ channels have been characterized, some of which have a medium-sized conductance, are TEA-sensitive, and are not blocked by apamin. A 110- to 125 pS channel described by [1] is sensitive to low concentrations of Ca2+, so that most of the channels are open at 0.1 μM, is activated by 30 mV depolarizations, and is blocked by charybdotoxin and by high concentrations of TEA. A 135 pS channel described by [6] is sensitive to submicromolar concentrations of Ca2+, has a Hill coefficient of ~ 2, has several closed states, has faster kinetics than the BK channel, and is blocked by charybdotoxin.
Because we did not have enough data available to describe the K2 channel completely, we used the same kinetic scheme as for the BK channel but with a lower voltage threshold, faster kinetics, and a lower half-activation Ca2+ concentration of 0.2 μM. Thus, despite the similar form of the equations, the K2 channel is quite different from the KC channel in voltage- and Ca2+ dependence.
[1] J Farley and B Rudy. Multiple types of voltage-dependent Ca2+-activated K+ channels of large conductance in rat brain synaptosomal membranes. Biophysical Journal, 53:919–934, 1988.
[2] DL Gruol, T Jacquin, and AJ Yool. Single-channel K+ currents recorded from the somatic and dendritic regions of cerebellar purkinje neurons in culture. Journal of Neuroscience, 11:1002–1015, 1991.
[3] B Lancaster, RA Nicoll, and DJ Perkel. Calcium activates two types of potassium channels in rat hippocampal neurons in culture. Journal of Neuroscience, 11:23–30, 1991.
[4] DG Lang and AK Ritchie. Large and small conductance calcium-activated potassium channels in the gh3 anterior pituitary cell line. Pfluegers Archiv, 410:614–622, 1987.
[5] R Latorre, A Oberhauser, P Labarca, and O Alvarez. Varieties of calcium-activated potassium channels. Annual Review of Physiology, 51:385–399, 1989.
[6] PH Reinhart, S Chung, and IB Levitan. A family of calcium-dependent potassium channels from rat brain. Neuron, 2:1031–1041, 1989.