![1
RT Mn(t) = --1----------------2+----
RMn + ΣGn(V, [Ca ],t)](pub-purkinje-deschutter1-discretization0x.png)
Related Documentation:
All simulations were performed with a model using the morphology of cell I of Rapp et al. (1994) unless otherwise noted.
Because electrotonic lengths were short, more computationally efficient asymmetric compartments (this GENESIS object corresponds to a three-element segment, as described by [4]) could be used. Simulations done with and without asymmetric compartments confirmed that the difference in input resistance (RN) and system time constant (τ0) for a passive membrane model was < 1 %.
In an active membrane compartment the total Rm is variable, resulting in a continuously changing space constant and electrotonic length [1, 3]. The total Rm (RTMn) of a compartment n at a specific time was computed as the sum of all conductances [G(V, [Ca2+],t) from Eqn. (1) and the passive component, RM n, determined by Rm
|
| (1) |
The compartment’s space constant λn could be derived from RTMn, its axial resistance RIn, and its length Ln
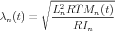 | (2) |
The compartment’s electrotonic length was the ratio of its length, Ln, over the space constant, λn. The electronic distance between two compartments was computed as the sum of the electronic lengths of all intervening compartments.
For those simulations that related to granule cell input, the total membrane surface was kept constant by subtracting the membrane surface of modeled spines from the membrane surface of the dendritic compartment to which they were connected. For a spiny dendritic compartment with length L and diameter D, the membrane surface (S in μm2), used for the computation of compartment membrane capacitance and resistance [2, 3], depended thus also on the number of simulated spines (Ns) connected to that compartment
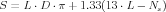 | (3) |
[1] Martin K A C Bernander Ö, Douglas R J and Koch C. Synaptic background activity influences spatiotemporal integration in single pyramidal cells. Proceedings of the National Academy of Sciences, 88:11569–11573, 1991.
[2] WR Holmes and CD Woody. Effects of uniform and non-uniform synaptic ‘activation-distributions’ on cable properties of modeled cortical pyramidal neurons. Brain Research, 505:12–22, 1989.
[3] M Rapp, Y Yarom, and I Segev. The impact of parallel fiber background activity on the cable properties of cerebellar Purkinje cells. Neural Computation, 4:518–533, 1992.
[4] I Segev, JW Fleshman, JP Miller, and B Bunow. Modeling the electrical behavior of anatomically complex neurons using a network analysis program: Passive membrane. Biological Cybernetics, 53:27–40, 1985.